
BIOME.cc – Biome evolution and climate change
Capucine Lechartre, Haozhi Ma, Elena Plekhanova, Manuel Richard Popp, Gengchen Yang
2024 - 2028
Kooperation- Federal University of Goiás, Brazil
- Helmholtz Zentrum für Umweltforschung
- Michigan State University, East Lansing, MI, USA
- National Aeronautics and Space Administration NASA
- Ohio State University, Columbus, OH, USA
- Peking University, Beijing, China
- University of Otago, Neuseeland
- University of Calgary, Canada
- USDA Forest Service, Missoula, MT, USA
Terrestrische Ökosysteme zeigen zunehmende Stresssymptome als Reaktion auf den fortschreitenden Klimawandel. In diesem Projekt kombinieren wir eine globale Analyse von Ökosystem-Störungen mit einer neuen Biomtheorie, um genauer vorherzusagen, wo auf der Erde welche Arten von Störungen als Reaktion auf den fortschreitenden Klimawandel auftreten können. Die Erprobung einer neuen Biomtheorie wird zudem ein besseres Verständnis dafür ermöglichen, wie sich Biome entwickelt haben und wie die globale Biodiversität entstanden und räumlich strukturiert wurde.
Globale Biome ¶
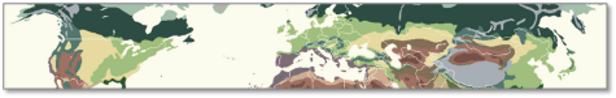
Der fortschreitende Klimawandel verändert die Zusammensetzung und die räumliche Vertbreitung der Biodiversität stetig. Wir wissen relativ gut, wie einzelne Arten auf den Klimawandel reagieren, verstehen aber sehr viel weniger gut, wie ganze Ökosysteme und globale Vegetationszonen (Biome) auf den Klimawandel reagieren. Globale Biome stellen strukturell und physiognomisch einheitliche Vegetationszonen dar, die sich auch klimatisch gut voneinander trennen lassen, wie etwa der „tropische Regenwald“, der „boreale Nadelwald“ oder die „Tundra“ (Abb. 1). Die strukturelle Einheitlichkeit von Biomen Einheitlichkeit ist ein Ergebnis davon, dass die dominanten Pflanzenarten von Biomen unabhängig von ihrer evolutiven Abstammung unter ähnlichem Klima überall auf der Erde sehr ähnliche Formen (Pflanzenmerkmale) entwickelt haben. Diese biomtypischen funktionellen Eigenschaften stehen für eine optimale Anpassung an das vorherrschende Klima und ermöglichen diesen Pflanzen, unter diesen Bedingungen zu dominieren. Ein Biom bedeckt eine relativ große Fläche auf der Erde und ist in der Regel auf mehreren Kontinenten zu finden. Innerhalb eines Bioms variiert das Klima und damit auch die Artenzusammensetzung; doch solange die klimatische Biom-Grenzen nicht überschritten wird, weisen die dominanten Pflanzenarten immer dieselben strukturellen und funktionellen Merkmale auf. Diese Eigenschaften sind das Ergebnis der Evolution der Pflanzen über Millionen von Jahren. Die Arten der tropischen Regenwälder in Südamerika, Afrika und Asien sind kaum miteinander verwandt, zu unterschiedlich sind ihre Entwicklungswege und ihre Abstammung. Und doch hat die Evolution der Pflanzen in allen drei Regionen unter ähnlichen Klimabedingungen zu sehr ähnlichen Strukturen geführt; wir nennen das konvergente Evolution.

Ziele des Forschungsprojekts ¶
Für dieses Projekt verfolgen wir zwei voneinander abhängig Ziele. Das erste Ziel zielt darauf ab, eine neue Biomtheorie zu erforschen und zu testen: die Biomgrösse-Stressresistenz-Theorie. Diese Theorie verspricht ein besseres Verständnis dafür, wie sich Biome durch evolutionäre Prozesse entwickelt haben und wie dies zur aktuellen Struktur und Verteilung der globalen Artenvielfalt geführt hat. Mit dem zweiten Ziel zielt beabsichtigen wir, Ökosystemstörungen als Reaktion auf den aktuellen und zukünftigen Klimawandel im Zusammenhang mit Biomverschiebungen zu bewerten und vorherzusagen.
1) Die Biomgrösse-Stressresistenz-Theorie ¶
Biome haben sich im Laufe der Jahrmillionen konvergenter Evolution unter ähnlichen klimatischen Bedingungen auf verschiedenen Kontinenten entwickelt. Die Flächen der Biome auf den Kontinenten sind jedoch nicht identisch, weder räumlich noch zeitlich. Dies bedeutet, dass ein Biom im Laufe der Zeit immer eine andere und sich verändernde Fläche für die Pflanzenevolution zur Verfügung hatte. Ausserdem kann es auf einem Kontinent verschiedene Flächen desselben Bioms gegeben haben, welche aber nicht immer gleich gut miteinander verbunden waren. Die Evolutionsbiologie hat die „Theorie des Biom-Konservativität“ formuliert, die davon ausgeht, dass Biome sog. Evolutionsarenen darstellen, in denen die Evolution mehr oder weniger räumlich eingeschränkt war, und dass die meisten neu-entwickelten Tochter-Arten im selben Biom verblieben sind. Wenn dies zutrifft, dann haben die unterschiedlichen räumlichen Dimensionen und die Konnektivität der Biomflächen im Laufe der Zeit zu unterschiedlicher Artenvielfalt pro Biom und Kontinent geführt, so wie es die „Theorie der Inselbiogeographie“ vorhersagt. Und wenn dies zutrifft, dann kann erwartet werden, dass unterschiedliche Artenvielfalt zwischen Flächen desselben Bioms zu Fitnessunterschieden führt, abiotischem und biotischem Stress zu widerstehen.
Daher formulieren wir hier die „Biom-Grösse – Stressresistenz-Theorie“, die vorhersagt, dass eine Biomfläche mit größerer Artenvielfalt eine höhere Toleranz der biomdominanten Arten gegenüber abiotischem (z. B. Kälte oder Dürre) oder biotischem (z. B. biologische Invasionen) Stress aufweist als eine andere Biom-Flöche mit geringere Artenvielfalt. In der Praxis bedeutet dies, dass ein Biom, das eine kleine Fläche auf dem Kontinent „A“ einnimmt, weniger tolerant gegenüber abiotischem oder biotischem Stress sein wird, als wenn dasselbe Biom eine grosse Fläche auf dem Kontinent „B“ einnimmt. Plausible Beispiele sind die hohen Raten biologischer Invasionen (biotischer Stress) auf kleinen und/oder lange getrennten Inseln (Hawai‘i, Neuseeland) oder Kontinenten (Australien) oder die Temperatur an der Baumgrenze auf kontinentalen (hohe Konnektivität) im Vergleich zu ozeanischen (geringe Konnektivität) Inseln (Abb. 2). Und der Begriff „Fläche“ steht für das Integral aus Fläche und Konnektivität, jetzt und in den letzten 5–10 (–20) Millionen Jahren, die für die Entwicklung der gegenwärtigen Pflanzenbiodiversität relevant sind. Um diese Theorie zu testen, formulieren wir mehrere Analyseschritte.

Analyseschritte
- Definition der globalen Biome anhand funktionaler und struktureller Eigenschaften ohne Verwendung von Klimaeigenschaften
- Kartierung globaler Biome anhand funktionaler und struktureller Eigenschaften
- Testen der “Biom-Konservativität” Theorie für die unterschiedlichen Biome der Welt
- Bewertung der Fläche und Konnektivität der historischen Biomflächen (letzte 10-20 Millionen Jahre mittels BIOME4)
- Bewertung der Evolution der Pflanzenvielfalt mit oder ohne Annahme des Biom-Konservativität (+/- strikte evolutive Grenzen zwischen Biomen) für die letzten 10-20 Millionen Jahre mit Gen3sis und Vergleich der Simulationsergebnisse mit biomspezifischen Pflanzen-Artenlisten, zusammengestellt aus globalen Datenbanken und Modellen
- Bestimmen der Klimagrenzen kartierter Biomflächen auf unterschiedlichen Kontinenten (aus Schritt 2), um die formulierte Theorie zu testen.
Derzeit definieren (1) und kartieren (2) wir Biome weltweit und beginnen mit der Analyse der Theorie des Biomkonservativismus für verschiedene Biome, indem wir sowohl Daten aus der vorhandene Literatur zusammenstellen als auch Phylogenien biom-dominanter Pflanzengruppen analysieren (3). Darüber hinaus beginnen wir mit der Erforschung der Flächegrösse und Konnektivität von Biomen mithilfe des dynamischen Prozessmodells BIOME4 über die letzten 10-20 Millionen Jahre (4). Basierend auf diesen Informationen werden wir später verschiedene Szenarien der Pflanzenevolution simulieren, indem wir mehr oder weniger strikte evolutive Grenzen zwischen Biomflächen über die letzten 10-20 Millionen Jahre postulieren (5). In diesem Zeitraum hat sich die Mehrheit der heute noch existierenden Pflanzenarten entwickelt und ist daher für die Prüfung unserer „Biom-Grösse – Stress-Resistenz“ Theorie relevant. Klimagrenzen der kartierten Biome werden im Zusammenhang mit dem zweiten Ziel identifiziert (6).
2) Ökosystem-Störungen aufgrund von potenzieller Biomverschiebung durch Klimawandel ¶
In den letzten 40 Jahren haben wir einen dramatischen Anstieg der globalen Temperaturen und damit verbundene regionale Veränderungen der Niederschlagsmuster erlebt. Während diese Veränderungen die potenziellen geografischen Grenzen aller großen Biome der Erde verschoben haben, hinken die tatsächlichen Verschiebungen der Biome erheblich hinterher. Das liegt daran, dass die Verschiebung Tausender Pflanzen, Tiere, Pilze und Mikroben Jahrzehnte, Jahrhunderte und Jahrtausende dauert. Daher gibt es viele Regionen mit einzelnen Biomfleächen, die nicht mehr im Einklang mit ihrem Klima sind (Abb. 3). Dies kann zu schweren abiotischen (Dürre, Feuer, Windwurf) oder biotischen (Schädlingsbefall, biologische Invasionen) Störungen führen. Wir können ausserdem davon ausgehen, dass die meisten Biome grundsätzlich mit Störungen umzugehen können, da dies Teil der Anpassung an die vorherrschenden Umweltbedingungen ist. Wenn jedoch der fortschreitende Klimawandel einen Teil des früheren Biomgebiets aufgrund des sich seit 1975 rasch ändernden Klimas aus dem Takt geraten lässt, können wir davon ausgehen, dass der Teil des Bioms, der nicht mehr dem geeigneten Biomklima ausgesetzt ist, Störungen erfährt, die schwerer und häufiger sind als in der Vergangenheit. Das Gleiche gilt für die Art und Weise, wie biom-typische Arten auf sich veränderndes Klima reagieren. Eine mögliche Biomverschiebung führt jedoch in der Regel dazu, dass andere funktionellen Pflanzenmerkmale für biomdominante Arten bevorzugt werden. Biomverschiebungen stellen daher eine massivere Fehlanpassung dar, als wenn der Klimawandel einen Wechsel von einer Art zu einer anderen nahelegt, wobei beide Arten die gleichen funktionellen Eigenschaften aufweisen.
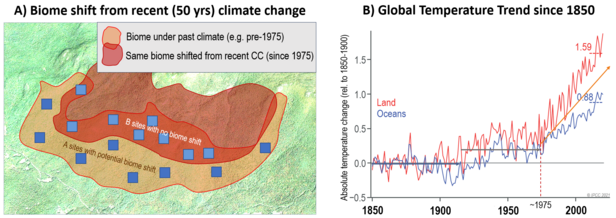
Daher stellen wir die Hypothese auf, dass „Gebiete mit einer kürzlich erfolgten potenziellen Biomverschiebung aufgrund des anhaltenden raschen Klimawandels stärkere und häufigere Störungen durch abiotischen und biotischen Stress aufweisen als benachbarte Regionen innerhalb desselben Bioms ohne potenzielle Biomverschiebung“ (Abb. 3). Störungen können durch verringerte oder erhöhte Niederschläge entstehen, mit oder ohne Kombination mit allgemein steigenden Temperaturen. Dies kann den Dürrestress im Allgemeinen verstärken, mit schwereren und längeren Dürreperioden, die zu Waldsterben, Wald- und Weidebränden oder Schädlingsbefall durch geschwächte dominante Arten führen. Dies kann jedoch auch biologische Invasionen verstärken, wo die Erwärmung nun andere funktionelle Pflanzentypen begünstigt, wie z. B. die Verschiebung der Baumgrenzen nach oben oder die Ausbreitung immergrüner Arten in die Laubwaldvegetation (sogenannte „Laurophilisierung“). In diesem Projekt werden wir diese Hypothese „potenzielle Biomverschiebung – Stressstärke/-häufigkeit“ für verschiedene Biomverschiebungen testen. Dies wird es uns ermöglichen, ein allgemeineres Prognosemodell zu formulieren, um vorherzusagen, welche Arten von Störungen wir angesichts des fortschreitenden Klimawandels für die verschiedenen Biome weltweit erwarten können.
Analyseschritte
- Verwenden der kontinentalen Klimagrenzen aus Teil 1f und Projizieren der potenzielle Biomverschiebung für die letzten 50 Jahre, um potenzielle Verschiebungsbereiche als Kontrast zu den Bereichen pro Biom abzubilden, die unverändert bleiben (siehe Abb. 3).
- Entwerfen eines Probenahmeprotokoll, um Störungen auf Biomebene durch Fernerkundung für Regionen mit potenzieller Biomverschiebung und nahe gelegene Bereiche ohne Verschiebung als Kontrast zu bewerten.
- Zuweisen der wahrscheinlichen Störungsart anhand von Fernerundungs-Analysen, Literaturrecherchen und Feldarbeit.
- Testen der Hypothese, ob es einen Störungsunterschied zwischen Regionen mit/ohne potenzielle Biome-Verschiebung gibt.
- Verallgemeinern der Störungsmuster durch potenzielle Biomverschiebungen auf zukünftige Bedingungen und Vorhersagen, wo und in welchem Biom wir welche Störung in Zukunft erwarten werden.
- Detaillierte Anlysen in drei warm- und kaltgemässigte (sowohl immergrünen als auch laubabwerfenden) Waldbiomen, einschließlich intensiverer Feldarbeit, um Störungsprozesse detaillierter zu untersuchen.
- Vergleich der Störungsunterschiede pro Biom weltweit, indem Störungen auf kontinentaler Ebene mit den Unterschieden in Artenvielfalt und Fläche dieser Biome pro Kontinent in Beziehung gesetzt werden.